
 Sometimes too much of a good thing can be a bad thing. Since their introduction, antibiotics have revolutionized modern medicine and saved countless lives. Yet, antibiotic use is the number one risk factor for the development of Clostridium difficile infections or CDIs.
Sometimes too much of a good thing can be a bad thing. Since their introduction, antibiotics have revolutionized modern medicine and saved countless lives. Yet, antibiotic use is the number one risk factor for the development of Clostridium difficile infections or CDIs.
C. difficile is an opportunistic bacterial pathogen that has emerged as the leading cause of health care associated infective diarrhoea (Slayton et al. 2015). Despite asymptomatic carriage of C. difficile among healthy individuals, antibiotic-induced disruption of the gut microbiota can create an environment permissive to C. difficile germination and growth (Smits et al. 2016). In toxigenic strains, germination of C. difficile spores into their toxin-producing vegetative state triggers a complex cascade of host cellular responses causing diarrhoea, inflammation, and tissue-necrosis (Smits et al. 2016). Clostridial toxin A (TcdA) and toxin B (TcdB) are large homologous toxins with similar multidomain architectures and represent primary determinants of C. difficile pathogenesis (Kuehne et al. 2014). As of late, several studies have shown that intestinal bile acids play important roles in modulating various aspects of the C. difficile lifestyle (Francis et al. 2013; Thanissery et al. 2017). In Tam et al., the authors unravel an entirely new role for bile acids in directly binding to and inhibiting C. difficile toxin function providing a framework for the development of novel anti-toxin therapeutics.
Building on their previous work, where they identified that a synthetic bile acid analog, methyl cholate, inhibited TcdB-induced toxicity of C. difficile, the authors set out to explore the biological and therapeutic significance of endogenous bile acids on toxin pathogenesis (Tam et al. 2015). To do this, they evaluated each of the individual human intestinal bile acids over a range of physiological relevant concentrations for their ability to interact with and modulate the activity of TcdA and TcdB. Strikingly, all the bile acids tested showed evidence of binding and inhibition of TcdB to varying extents, whereas none interacted with or inhibited TcdA. This is of particular interest given that TcdB has been shown to be the primary determinant of disease in humans. Moreover, the authors found that secondary bile acids, naturally produced by metabolization of primary bile acids by the microbiota, are more potent than their corresponding primary bile acid precursors, despite differing only by a single hydroxyl group. For example, taurolithocholic acid (TLCA) was more effective at inhibiting TcdB-induced cell rounding, a phenotype of toxin-mediated cellular toxicity, compared to its precursor, taurochenodeoxycholic acid (TCDCA). Nonetheless, the primary bile acid, TCDCA, was shown to protect human colonic cells from TcdB activity by preventing the disruption of monolayer integrity and transepithelial electrical resistance.
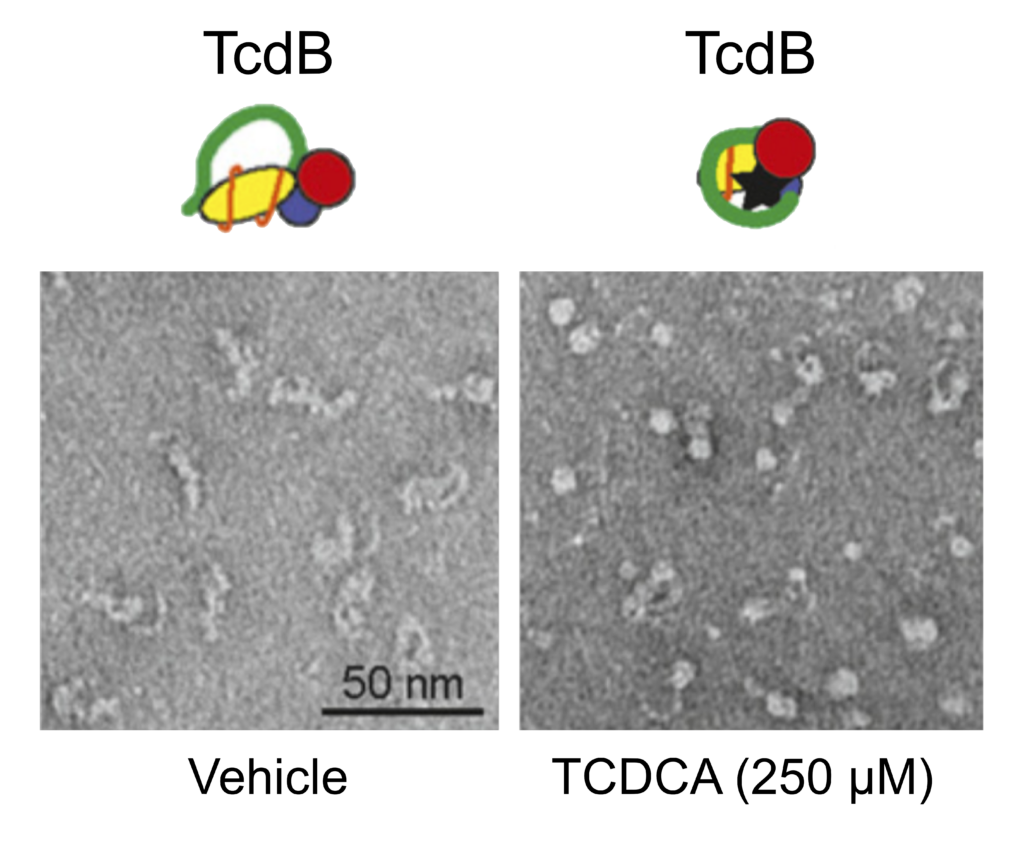
Given the disparity between bile acid activity against TcdA and TcdB, the authors focused their attention on the sequence divergence between these two toxins. They found the most divergent region to be within the C-terminal combined repetitive oligopeptide repeats (CROP) domain, where TcdB has fewer repeats than TcdA. To test the hypothesis that the bile acids may be directly binding to CROP, the authors first took advantage of the naturally “CROP-less” homolog TpeL from Clostridium perfringens. They observed no evidence of TCDCA binding to TpeL, supporting the possible role for CROP in binding bile acids. Next, the authors generated truncations within the TcdB toxin CROP domain and observed that removing as few as four terminal repeats rendered TcdB insensitive to the bile acid, TCDCA. Previous studies have reported that the C-terminal CROP domain of TcdB adopts a dynamic conformation relative to the main body of the toxin. Thus, the authors tested the ability of bile acid binding to modify the conformational structure of TcdB upon binding. Using negative-stain electron microscopy, Tam et al. showed that TCDCA and the synthetic inhibitor, methyl cholate was able to induce TcdB to adopt an increasingly “balled up” conformation resulting in reduced overall size and protection from proteolytic degradation. Given the dramatic changes in TcdB structure, the authors investigated the ability of TcdB to bind the cell surface receptors, the first step in the intoxication pathway of TcdB. In the absence of bile acids, TcdB was recovered from the cell surface of human colonic cells, however, preincubation with TCDCA completely blocked cell surface binding.
Notwithstanding the ability of bile acids to provide protection against TcdB-induced toxicity to human cells, bile acids themselves part take in a variety of important physiological functions, and are therefore, unlikely therapeutic agents. To identify other potentially more “drug-like” molecules that could serve as novel antitoxins, the authors conducted a high-throughput screen for compounds that bind to TcdB and protect cells from TcdB-induced toxicity. This screen identified ethaverine, a previously approved peripheral coronary vasodilator drug, and demonstrated that it is capable of inhibiting TcdB in a bile acid-like mechanism.
Together, this work identifies an unexpected role for bile acids in binding to and inhibiting the function of TcdB via a CROP-dependent mechanism of action. Identification of ethaverine as a novel inhibitor of TcdB-mediated toxicity demonstrates the potential to exploit this unique mechanism of action for the development of therapeutic inhibitors of C. difficile pathogenesis.
Primary Reference:
Tam, J. et al. Intestinal bile acids directly modulate the structure and function of C. difficile TcdB toxin. PNAS 117, (2020).
References:
Slayton, R.B. et al. The cost-benefit of federal investment in preventing Clostridium difficile infections through the use of a multifaceted infection control and antimicrobial stewardship program. Infect. Control Hosp. Epidemiol. 36, (2015).
Smits, W.K., Lyras, D., Lacy, D.B., Wilcox, M.H. & Kuijper, E.J. Clostridium difficile infection. Nat. Rev. Dis. Primers 2, (2016).
Kuehne, S.A. et al. Importance of toxin A, toxin B, and CDT in virulence of an epidemic Clostridium difficile strain. J. Infect. Dis. 209, (2014).
Francis, M.B., Allen, C.A., Shrestha, R. & Sorg, J.A. Bile acid recognition by the Clostridium difficile germinant receptor, CspC, is important for establishing infection. PLoS Pathog. 9, (2018).
Thanissery, R., Winston, J.A. & Theriot, C.M. Inhibition of spore germination, growth, and toxin activity of clinically relevant C. difficile strains by gut microbiota derived secondary bile acids. Anaerobe 45, (2017).
Tam, J. et al. Small molecule inhibitors of Clostridium difficile toxin B-induced cellular damage. Chem. Biol. 22, (2015).

Cowen Lab


